Jasmonic acid

Jasmonic acid (Japanese Agricultural Standards mon さんるい British: jasmonate, JA) and the relative are lipid hormone signal matter controlling the change of the wide plant from growth and photosynthesis to reproduction outbreak. Particularly, JA is important to plant defense () and a poor environmental condition and other non-biology, the reply of the plant for the biologic stress to eating [1]. The plant releases JA as volatile organic compounds (VOC) to recall me to talking trees appearing in the fiction and can take the communication to be able to take the preparation to danger between plants [2].
In 1962, jasmonic acid methyl was isolated from jasmine oil derived from jasmine Jasminum grandiflorum, and structure of jasmonic acid became clear [3].
Table of contents
Structure of jasmonic acid and the relative





Jasmonic acid (JA) oxy; is composed in the chloroplast film which is a re-pin (oxygenicity fatty acid) by existing α-linolenic acid. The composition is started by conversion to 12-oxo - フィトジエン acid (OPDA) of the α-linolenic acid;, (12-オキソフィトジエン acid reductase), is (+) by the oxidation of reduction and three phases next -7-iso-JA is formed. All reactions after the beginning happen only for conversion from α-linolenic acid to OPDA with peroxisome in a chloroplast [4].
JA oneself is metabolized to an active form or an inactivated form relative more. Jasmonic acid methyl (MeJA) is the volatile compound which may participate in the communication between the plant. JA is combined with isoleucine (Ile) of the amino acid and becomes JA-Ile. (+) where JA-Ile is more unstable like JA -(+) where I am in equilibrium, but the bioactivity cries in JA-Ile with 7-iso-JA-Ile -Is JA relative necessary for the JA signal transmission that 7-iso-JA-Ile is known to alone now [4]; [5].
Jasmonate biosynthesis

The biosynthesis course of jasmonic acid was clarified by Brady Vick and Don Zimmerman [6]; [7]. At first α-linolenic acid (1) is 13-(S) by a 13-lipoxygenase (13-LOX, A) -It is oxidized to hydroperoxy linolenic acid (2). Epoxidation is beginning, 12,13-(S) by 13-arene oxide synthetase (13-AOS, B) next -Epoxy -9(Z),11(E),15(Z) -It becomes octadecatrienoic acid (3). 12-oxo - フィトジエン acid (OPDA, 4) is formed by arene oxide cyclase (AOC, C) successively. It is (+) by beta-oxidation of reduction with 12-oxo - フィトジエン acid reductase and 3 degrees more -7-iso- jasmonic acid (5a) is formed and is the diastereomer which is thermodynamically stable by the last (-) -Becoming it takes place Eppie to jasmonic acid (5b).
Jasmonic acid signal transmission mechanism
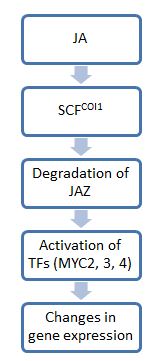
Generally, I closely resemble a thing of the auxin signal transmission for the stages in jasmonic acid (JA) signal course. It is from the E3 ubiquitin ligase complex which couples ubiquitin to a substrate so that it is disintegrated for the first stage by proteasome. The second uses a transcription factor influencing a physiological change. One of the key molecules in this course is JAZ protein working as an ON/OFF switch of the JA signal transmission. When JA lacks, the JAZ protein family binds to the downstream transcription factor and limits those activity. However, the JAZ is broken down in the presence of JA or the bioactive relative, and a transcription factor concerned with gene expression necessary for a stress reply is isolated [8].
Because the resolution of the JAZ does not happen with the null coi1 mutant plant, it is thought that COI1 protein transmits the resolution of the JAZ. COI1 belongs to F-box protein () stored highly and leads a substrate to E3 ubiquitin ligase () SCFCOI1. This complex which is finally formed is known as an SCF complex () [9]. These complexes are combined with JAZ and aim for the resolution with the proteasome. However, all JA relatives do not activate this signal course when I think about a wide spectrum of the JA molecules group, and the range of JA molecules participating in this course is unidentified [4]. It is shown until now that only 7-iso-JA-Ile is necessary for the resolution of JAZ11 which COI1 transmits [5]. 7-iso-JA-Ile and the relative similar structurally can bind to the COI1-JAZ complex and promote ubiquitin and the latter resolution to thereby have [4].
The possibility that COI1 acts as a receptor in the cells of the JA signal by this mechanism model increases. In the recent study, this hypothesis was supported by it having been found that it acted as coexistence receptor (co-receptor) where COI1-JAZ complex recognized JA. 7-iso-JA-Ile is connected to be concrete in the both sides of 20 amino acid parts of the saved Jas motif in ligand-binding pockets in COI1 and JAZ. This JAZ residue acts as a stopper of the pockets of COI1 and keeps 7-iso-JA-Ile the state that bound to the pocket. In addition, by the corefinement with COI1 and inositol 5 phosphoric acid () (InsP5) and a removal experiment of subsequent InsP5, InsP5 is the element which is essential for a coexistence receptor, and strengthen a coexistence receptor complex; it is suggested to play a role [10].
The transcription factor activates a gene necessary for specific JA reply when freed from JAZ. The transcription factor studied most acting by this course belongs to the MYC family. Basic helix-loop-helix (bHLH) DNA-binding motif features the MYC family. These factors (MYC2, 3, 4) tend to act additively. For example, the plant which suffered a loss only in one myc comes to show sensitivity for the contact of the insect than a normal plant more. The plant which suffered a loss in all three myc becomes the sensitivity for an injury like coil1 mutant. This plant does not show JA responsiveness at all and cannot start defense for the contact. These MYC molecules share a function, but these expression styles and transcription functions are greatly different. For example, MYC2 has a big effect by the extension of the root in comparison with MYC3 and MYC4 [11].
In addition, a loop goes back, and MYC2 controls an expression of JAZ level (negative feedback loop); [11]. All these transcription factors have different influence in a JAZ level after the JA signal transmission. The JAZ level influences a transcription factor and a gene expression level equally.
Role of jasmonic acid
Jasmonic acid (JA) controls many different processes in a plant, but roles in the replies to injury are understood best. Following a mechanical injury or eating, JA biosynthesis is activated immediately, and an appropriate reply gene emerges. For example, with the tomato, defense molecules obstructing the digestion of the leaf in the bowels of the insect are produced by an injury. An indirect result of the JA signal transmission includes release for the volatilization of the compound coming from JA. MeJA which there is on a leaf moves to a plant nearby through the air and causes the protective reaction for the injury to the plant which is not damaged [1].
Following a role in the defense, JA is related to cell death and the aging of the leaf. JA interacts with much kinase and transcription factors about aging. JA induces the cell death due to mitochondria by deriving accumulation of active oxygen (ROS) again. These compounds ruin a mitochondrial film, and they cause apoptosis (apoptosis). It is thought that the plant prevents infection from spreading further by inducing cell death by oneself [12].
JA and the relative participate in outbreak, the symbiosis of the plant and the following processes again.
- It was found that JA obstructed the extension of the root by the early study of the JA overexpression mutant. The mechanism of this phenomenon has not been yet clarified, but it is found that COI1 course is essential in the tensile inhibition of the root for some reason with the mutant of the COI1-dependent signal transmission course because a tendency to do a phenomenon this inhibition is shown [11]; [13].
- JA plays many roles in the outbreak of the flower. Mutants of JA composition of Arabidopsis or the JA signal transmission usually show male sterility by a delay of the development. Interestingly, the same gene promoting male sterility in Arabidopsis promotes femininity sterility with the tomato. The overexpression of 12-OH-JA also causes a delay of the flowering [13].
- JA and MeJA obstruct the germination of the non-dormancy seed and stimulate the germination of the dormancy seed [14].
- High-level JA promotes the accumulation of the deposit protein. The gene encoding a plant deposit protein is JA-responsive. Specifically, the ツベロン acid (12-hydroxy-epi-jasmonic acid, tuberonic acid, TA) of the JA relative derives the formation of tuber (tuber) [15]; [16].
- JA plays its part in a plant and the compulsion with the microbe. However, the accurate role is still unknown. It is thought that JA controls a legume and signal exchange and the root nodule form straight control between the root nodule bacteria now. On the other hand, it is thought that the rise in JA level controls distribution and the stress tolerance of carbohydrates in a mycorrhiza plant [17].
- As for 12-hydroxy jasmonic acid glucoside, COI1-JAZ course controls the nyctinasty of the American silk tree (Samanea saman) independently [18]; [19].
Jasmonic acid and etiology
Pseudomonas syringae causes bacterial spot blotch in a tomato by taking over jasmonic acid (JA) signal transmission course of the plant. These bacteria pour cocktail of the viral effector protein into a host cell using a type III secretion device.
To this mixture, one of the molecules to be included in is the corona Japanese spaniel of the plant toxin (coronatine, COR). (+) where the corona Japanese spaniel is active form JA -I resemble 7-iso-JA-Ile structurally. The JA non-sensitivity plant shows high resistance in P. syringae and does not reply to COR. Furthermore, I can regain the infectiousness of COR loss bacteria when I add MeJA. The plant infected with develops downstream JA and an injury reply gene, but the infection-specific genetic level is controlled. These data suggest that COR acts through JA course and erodes with a host plant. A hypothesis to sacrifice the defense for disease-causing germs is appointed the activation of the injury reply as. When JA injury reply course is activated, P. syringae can dodge an attack from the immune system of the host and can be infected more effectively [20].
Other crosstalk with the defense course
The jasmonic acid (JA) course is important to an injury reply, but is not the only signal transmission course to introduce the defense of the plant into. I do a specific reply for a non-creature and a biologic attack to build effective defense at the same time though it is most suitable, and there must be crosstalk between the defense courses that are different to fine-tune it again.
One studied most in JA in question is crosstalk with the salicylic acid (SA). The SA that is plant hormone mediates the defense for disease-causing germs by guiding infection-specific genetic expression and whole body acquisition resistance (systemic acquired resistance, SAR) together. SAR is that the plant which caught the attack of disease-causing germs locally gets the resistance for disease-causing germs with a whole body.
I can see an injury and the disease-causing germs reply to interact adversely each other. For example, the SAR decreases, but the eating resistance for the insect increases when I perform silencing of phenylalanine ammonia lyase (PAL) which is an enzyme composing an SA precursor. Similarly, the overexpression of the PAL raises SAR, but the injury reply after insect eating decreases [21]. Generally, the disease-causing germs surviving in a straight plant cell show sensitivity for SA instruction type defense more, but it is found that a phytophagous insect receiving a benefit from cell death and the disease-causing germs show sensitivity for JA instruction type defense more. Thus, this trade-off to be able to put on the defense course optimizes defense and protects plant resources [22].
The crosstalk happens between other plant hormone courses such as JA and abscisic acid (ABA) and ethylene (ET). These interaction optimizes the defense for the disease-causing germs with the different lifestyle and the grazing animal equally. For example, the MYC2 activity is stimulated by JA and both ABA courses and can unify the signals from both courses. Other transcription factors such as ERF1 call JA, and they happen as a result of ET signal transmission. All these molecules share it to activate a specific injury reply gene and work [22].
Finally the crosstalk is not limited to defense. The interaction of JA and ET is important in the outbreak, and the balance between these 2 molecules is indispensable to normal tip key to Arabidopsis seedling form projection outbreak. A further study is necessary to elucidate molecules controlling such a crosstalk [21].
Footnote
- ^ a b Farmer, E. E.; Ryan, C. A. (1990). "Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves". Proc. Natl. Acad. Sci. USA 87 (19): 7713–7716. doi: 10.1073/pnas.87.19.7713. PMC 54818. It is . PMID 11607107
- ^ Baldwin, I. T.; Halitschke, R., Paschold, A., von Dahl, C. C., Preston, C. A. (2006). "Volatile signaling in plant-plant interactions: "talking trees" in the genomics era." It is 812–815. doi: Science 311 10.1126/science.1118446. PMID 16469918.
- ^ Demole E; Lederer, E.; Mercier, D. (1962). "Isolement et détermination de la structure du jasmonate de méthyle, constituant odorant caractéristique de l'essence de jasmin." Helv. Chim. It is 675–685. doi: Acta 45 10.1002/hlca.19620450233.
- ^ a b c d Katsir, L.; Chung, H. S., Koo, A. J. K., Howe, G. A. (2008). "Jasmonate signaling: a conserved mechanism of hormone sensing". Curr. Opin. Plant Biol. 11: 428–435. doi: 10.1016/j.pbi.2008.05.004. PMC 2560989. It is . PMID 18583180
- ^ a b Fonseca S, Chini A, Hamberg M, Adie B, Porzel A, Kramell R, Miersch O, Wasternack C, Solano R (2009). "(+)-7-iso-Jasmonoyl-L-isoleucine is the endogenous bioactive jasmonate." Nat. Chem. Biol. 5 (5): 344-350. doi: 10.1038/nchembio.161. PMID 19349968.
- ^ Vick, BA. und Zimmerman, DC. (1984). "Biosynthesis of Jasmonic Acid by Several Plant Species." Plant Physiol. 75 (2): 458–461. doi: 10.1104/pp.75.2.458. PMID 16663643.
- ^ Vick, BA. und Zimmerman, DC. (1987). "Pathways of Fatty Acid Hydroperoxide Metabolism in Spinach Leaf Chloroplasts." Plant Physiol. 85 (4): 1073–1078. doi:10.It is 1073 4 1104/ pp. 85. PMID 16665806.
- ^ Chini, A.; Fonseca, S., Fernandez, G., Adie, B., Chico, J. M., Lorenzo, O., Garcia-Casado, G., Lopez-Vidriero, I., Lozano, F. M., Ponce, M. R., Micol, J. L, and Solano, R. (2007). "The JAZ family of repressors is the missing link in jasmonate signaling." Nature 448 (7154): 666–671. doi: 10.1038/nature06006. PMID 17637675.
- ^ Devoto, A; Nieto-Rostro, M, Xie, D, Ellis, C, Harmston, R, Patrick, E, Davis, J, Sherratt, L, Coleman, M, Turner, JG (2002). "COI1 links jasmonate signalling and fertility to the SCF ubiquitin-ligase complex in Arabidopsis." Plant J. 32 (4): 457–66. PMID 12445118.
- ^ Sheard, Laura B.; Tan, Xu, Mao, Haibin, Withers, John, Ben-Nissan, Gili, Hinds, Thomas R., Kobayashi, Yuichi, Hsu, Fong-Fu, Sharon, Michal, Browse, John, He, Sheng Yang, Rizo, Josep, Howe, Gregg A., Zheng, Ning (2010). "Jasmonate perception by inositol-phosphate-potentiated COI1–JAZ co-receptor". Nature 468 (7322): 400–405. doi: 10.1038/nature09430. PMC 2988090. It is . PMID 20927106
- ^ a b c Fernández-Calvo, P.; Chini, A., Fernández-Barbero, G., Chico, J.-M., Gimenez-Ibanez, S., Geerinck, J., Eeckhout, D., Schweizer, F., Godoy, M., Franco-Zorrilla, J. M., Pauwels, L., Witters, E., Puga, M. I., Paz-Ares, J., Goossens, A., Reymond, P., De Jaeger, G., Solano, R. (2011). "The Arabidopsis bHLH Transcription Factors MYC3 and MYC4 Are Targets of JAZ Repressors and Act Additively with MYC2 in the Activation of Jasmonate Responses". Plant Cell 23 (2): 701–715. doi: 10.1105/tpc.110.080788. PMC 3077776. It is . PMID 21335373
- ^ Reinbothe, C; Springer, A, Samol, I, Reinbothe, S (2009). "Plant oxylipins: role of jasmonic acid during programmed cell death, defence and leaf senescence." FEBS J. 276 (17): 4666–81. doi: 10.1111/j.1742-4658.2009.07193.x. PMID 19663906.
- ^ a b Wasternack, C. (2007). "Jasmonates: An Update on Biosynthesis, Signal Transduction and Action in Plant Stress Response, Growth and Development". Ann. Bot. 100 (4): 681–697. doi: 10.1093/aob/mcm079. PMC 2749622. It is . PMID 17513307
- ^ Creelman, Robert A.; Mullet, John E. (1997). "Biosynthesis and action of jasmonates in plants." Annu. Rev. Plant Physiol. Plant Mol. Biol. 48 (1): 355–381. doi: 10.1146/annurev.arplant.48.1.355. PMID 15012267.
- ^ Anderson, JM. (1988). "Jasmonic acid-dependent increases in the level of specific polypeptides in soybean suspension cultures and seedlings." J. Plant Growth Regul. 7: 203–11. doi: 10.1007/BF02025263.
- ^ Pelacho, AM; Mingo-Castel, AM. (1991). "Jasmonic acid induces tuberization of potato stolons cultured in vitro". Plant Physiol. 97: 1253–1255. doi: 10.1104/pp.97.3.1253. PMC 1081150. It is . PMID 16668517
- ^ Hause, Bettina; Schaarschmidt, Sara (2009). "The role of jasmonates in mutualistic symbioses between plants and soil-born microorganisms." Phytochemistry 70 (13-14): 1589–1599. doi: 10.1016/j.phytochem.2009.07.003. PMID 19700177.
- ^ Ueda, M.; Okazaki, M.; Ueda, K.; Yamamura, S. (2000). "A Leaf-Closing Substance of Albizzia julibrissin Durazz". Tetrahedron 56 (41): 8101–8105. doi: It is .
This article is taken from the Japanese Wikipedia Jasmonic acid
This article is distributed by cc-by-sa or GFDL license in accordance with the provisions of Wikipedia.
In addition, Tranpedia is simply not responsible for any show is only by translating the writings of foreign licenses that are compatible with CC-BY-SA license information.
0 개의 댓글:
댓글 쓰기